2 – Transporte de Gases Entre os Alvéolos e as Células
Após a ventilação dos alvéolos ocorre a difusão de oxigênio dos alvéolos para o sangue presente na circulação pulmonar e a difusão do dióxido de carbono na direção oposta. A difusão ocorre devido ao movimento das moléculas dos gases. A velocidade de difusão de cada um dos gases participantes da respiração é diretamente proporcional à pressão desse gás, é a chamada de pressão parcial do gás.
Cada gás contribui para a pressão total em proporção direta à sua concentração. Os gases dissolvidos na água e nos tecidos do corpo também exercem pressão, visto as moléculas dissolvidas estarem em movimento aleatório e terem energia cinética. Quando o ar penetra nas vias respiratórias, a água das superfícies dessas vias imediatamente se evapora e umedece o ar. Isto é resultado do fato de que as moléculas de água (vapor d'água), assim como as diferentes moléculas de gases dissolvidos no líquido, estão continuamente escapando da superfície de água.
A pressão que as moléculas de água exercem para escapar através da superfície é chamada de pressão de vapor da água. A difusão efetiva de um gás de área de alta pressão para área de baixa pressão é igual ao número de moléculas que se movimentam nesta direção menos o número que se movimenta na direção oposta, e isto, por sua vez, é proporcional à diferença entre a pressão de gás das duas áreas, é a chamada diferença de pressão de difusão.
Todos os gases que têm importância respiratória são altamente solúveis em lipídios e, conseqüentemente, altamente solúveis nas membranas celulares. O ar alveolar não tem a mesma concentração de gases que o ar atmosférico, devido ao fato de que a cada ciclo respiratório o ar alveolar é parcialmente renovado pelo ar atmosférico, o oxigênio está constantemente sendo absorvido do ar alveolar e o dióxido de carbono se difundindo do sangue pulmonar para os alvéolos.
À medida que entra nas vias respiratórias, o ar é exposto aos líquidos que revestem as superfícies respiratórias e é totalmente umidificado antes de entrar nos alvéolos. Somente 350 ml de ar fresco são trazidos para os alvéolos em cada inspiração normal e a mesma quantidade é eliminada a cada expiração, de modo que muitas inspirações são necessárias para substituir a maior parte do ar alveolar. Esta substituição lenta do ar alveolar é importante para impedir mudanças bruscas nas concentrações gasosas do sangue.
O oxigênio está constantemente sendo absorvido pelo sangue nos pulmões, e oxigênio novo, da atmosfera, está continuamente chegando aos alvéolos. Quanto mais rapidamente o oxigênio for absorvido, mais baixa será sua concentração nos alvéolos; por outro lado, quanto mais rápido o oxigênio novo é inspirado pelos alvéolos, mais alta será sua concentração. O dióxido de carbono é continuamente formado no organismo, sendo em seguida descarregado nos alvéolos e removido pela ventilação.
As concentrações e pressões de oxigênio e de dióxido de carbono nos alvéolos são determinadas pelas velocidades de absorção ou de excreção dos dois gases e também pelo nível de ventilação alveolar. A unidade respiratória é formada por um bronquíolo respiratório, ductos alveolares e alvéolos. As paredes destas estruturas possuem uma extensa rede de capilares interconectados, conhecida como membrana respiratória.
A partir de estudos histológicos estima-se que a superfície total da membrana respiratória tenha área de aproximadamente 50 a 100 metros quadrados no adulto normal. Os principais fatores que a determinam a velocidade da difusão gasosa através da membrana respiratória são a espessura da membrana, sua área superficial, a velocidade de difusão do gás e a diferença de pressão entre os dois lados da membrana.
Quando o sangue arterial alcança os tecidos periféricos, a pressão parcial do oxigênio é maior do que a pressão parcial de oxigênio no líquido intersticial. Essa diferença de pressão causa a rápida difusão do oxigênio, do sangue para os tecidos. Quando o oxigênio é utilizado pelas células, a maior parte dele é transformada em dióxido de carbono e este tem aumentada a sua pressão parcial intracelular. Como consequência, o dióxido de carbono se difunde das células para os capilares teciduais, sendo levado pelo sangue para os pulmões, onde se difunde dos capilares pulmonares para o espaço alveolar, e daí para fora do organismo. Normalmente, cerca de 97% do oxigênio transportado dos pulmões para os tecidos é carregado em combinação química com a hemoglobina nas hemácias, e os 3% restantes são transportados dissolvidos no plasma.
Assim, em condições normais, o oxigênio é transportado para os tecidos quase totalmente pela hemoglobina. Quando a pressão parcial de oxigênio está alta, como nos capilares pulmonares, o oxigênio de liga com a hemoglobina, mas quando a pressão parcial de oxigênio está baixa, como nos capilares teciduais, o oxigênio é liberado da hemoglobina. Esta é a base para quase todo o transporte de oxigênio dos pulmões para os tecidos.
Sob condições normais, a velocidade de utilização de oxigênio pelas células é controlada, em última análise, pela velocidade de consumo energético dentro das células, isto é, pela velocidade com que o ADP é produzido a partir do ATP.
O monóxido de carbono se combina com a hemoglobina no mesmo ponto onde o oxigênio se associa e, por conseguinte, pode deslocar o oxigênio da hemoglobina.
Além disso, ele se liga à hemoglobina com 250 vezes mais firmeza que o oxigênio. Um paciente gravemente envenenado com monóxido de carbono deve ser tratado administrando-se oxigênio puro, pois o oxigênio em altas pressões alveolares desloca o monóxido de carbono mais rapidamente do que o oxigênio de menor pressão parcial, que é encontrado no ar atmosférico.
O dióxido de carbono pode ser transportado sob a forma dissolvida no plasma (7%); ou combinando-se com a água no interior das hemácias para formar ácido carbônico e, em seguida os íons hidrogênio e bicarbonato (este processo ocorre dentro da hemácia e é catalizado pela enzima anidrase carbônica) (70%); e combinado com a hemoglobina (carboxiemoglobina) (15 a 25%). A formação de ácido carbônico pelo dióxido de carbono que passa dos tecidos para o interior das hemácias diminui o pH sanguíneo.
Contudo, a reação deste ácido com os tampões - (Os tampões são substâncias que limitam as variações do pH do sangue e demais líquidos orgânicos, ao se combinarem com os ácidos ou bases que alcançam esses líquidos. As substâncias que constituem os tampões agem aos pares ou, menos comumente, em grupos, constituindo um sistema protetor.
Um sistema tampão é constituído por um ácido fraco e o seu sal, formado com uma base forte. O ácido fraco e o sal do sistema tampão, em condições normais, existem em uma relação constante, que o organismo tende a preservar. Se gotejarmos continuamente ácido clorídrico em água durante um intervalo de 90 minutos, verificamos que o pH da água passa de 7 para 1,84. Se administrarmos proporcionalmente, a mesma quantidade de ácido clorídrico a um cão no mesmo período de tempo, verificamos que o pH do sangue do animal passa de 7,44 para 7,14. A diferença de comportamento diante da mistura com o ácido clorídrico reflete a atuação dos sistemas tampão do plasma do animal, que impedem a variação mais acentuada do pH.) - do sangue impede que a concentração de íons hidrogênio aumente muito (e que o pH desça muito). Normalmente, o sangue arterial tem um pH de aproximadamente 7,41 e, à medida que o sangue adquire dióxido de carbono nos capilares teciduais, o pH desce para um valor de aproximadamente 7,37. Ocorre o inverso quando o dióxido de carbono é liberado do sangue para os pulmões, com o pH se elevando para o valor arterial.
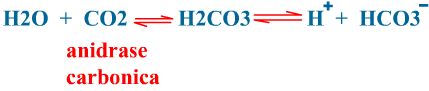


Nenhum comentário:
Postar um comentário